Control of gene expression requires factors that facilitate interactions between DNA, RNA, and protein. The Drosophila behavior/human splicing (DBHS) family of proteins mediate various protein-protein and protein-nucleic acid interactions, and influence gene expression in pathways as diverse as those relevant to neurobiology,2 development,3 innate immunity,4 cancer,5 and DNA repair.6
One member of the DBHS family of proteins, non-POU domain-containing octamer-binding protein (NONO), is emerging as central to the link between feeding and metabolism. In mice, NONO binds to pre-mRNA of genes involved in hepatic glucose and lipid metabolism, and post-transcriptionally regulates the rhythmic expression of these genes in response to the presence of nutrients.7 Using immunoprecipitation and immunostaining techniques, the number of target RNAs bound by NONO, as well as the number of NONO positive foci, were found to increase in hepatic cell nuclei of mice upon feeding.7 NONO was also identified as a component of paraspeckles, sub-nuclear bodies that regulate gene expression by retaining RNA in the nucleus.7 Furthermore, a large proportion of NONO-bound RNAs encode proteins involved in macronutrient metabolism and glucose uptake. In NONO-deficient mice, the rhythmic pattern of gene expression in response to nutrient availability is disrupted, resulting in reduced glucose tolerance as well as glycogen and lipid storage capabilities.7
These recent findings suggest that NONO post-transcriptionally coordinates RNA expression of genes involved in nutrient metabolism in response to nutritional stimuli. Identification of NONO as a novel player involved in the maintenance of metabolic homeostasis during the feeding-fasting cycle also suggests that you are not only what you eat, but also when you eat it!
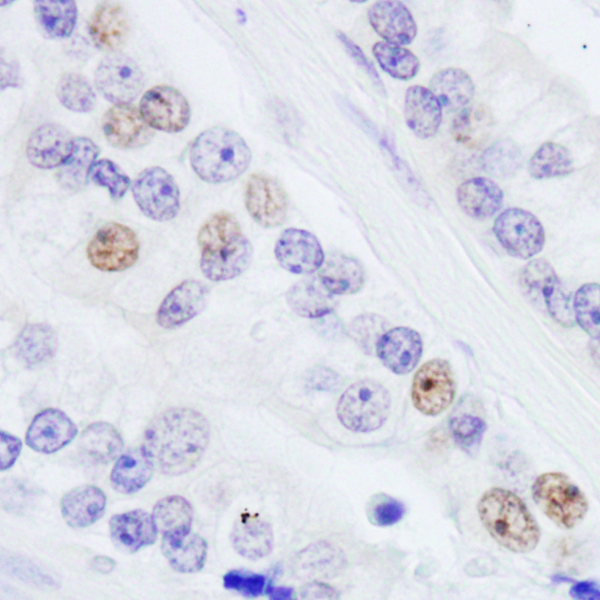
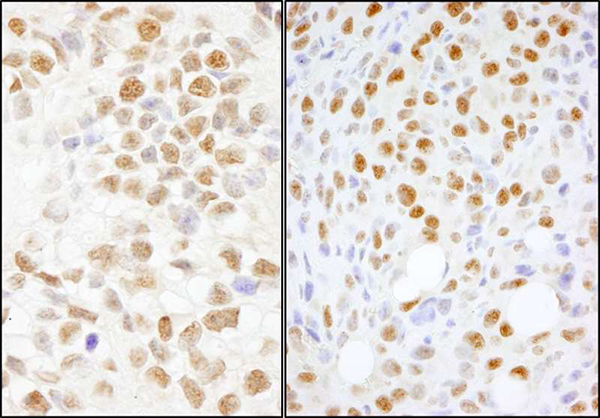
Bethyl offers the following NONO antibodies: NONO, NONO, NONO AbVantage™ Pack, NONO IHC