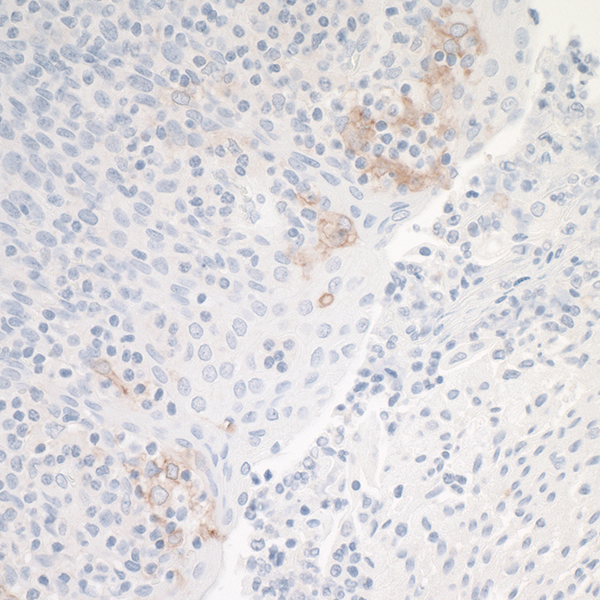
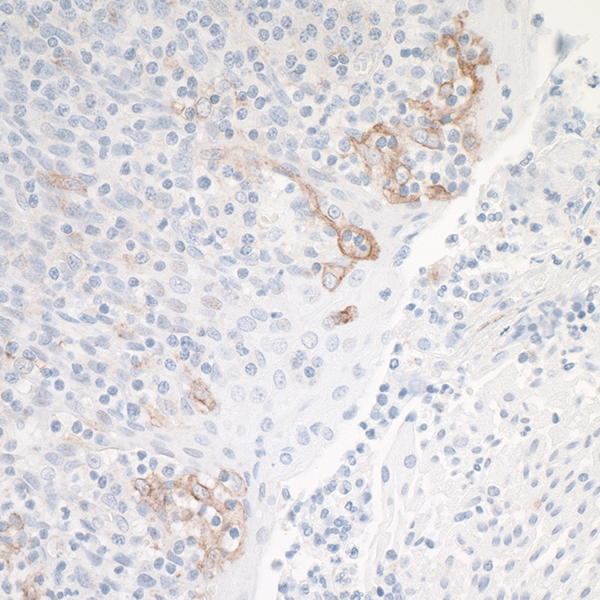
The most commonly known checkpoint that can be targeted therapeutically is the PD-1/PD-L1 pathway. In fact, FDA approval for several drugs targeting this pathway took place in 20172. PD-1 is a receptor found on T cells, and is upregulated on “exhausted” T cells that have seen chronic antigen stimulation3. Because tumors are an antigen-rich environment, tumor-infiltrating T cells, or TIL, often and rapidly begin to upregulate PD-1. Many tumor cells express PD-L1 on their cell surface4,5. Interaction between PD-L1 and PD-1 expressed on T cells leads to the transmission of a regulatory signal to the T cells6. Therefore, blocking the interaction between PD-1 and PD-L1 increases the effector function of TIL and lead to an anti-tumor immune response that extends progression-free and overall survival in a subset of cancer patients7.
Following the success of anti-PD-(L)1 therapy, several emerging checkpoint inhibitors have been tested in model systems. Signaling through these pathways confers a similar inhibitory effect on T cells to that downstream of PD-1. LAG-3, BTLA, TIM-3, and TIGIT are some of the receptors that T cells use to downregulate their response to antigen and have been proposed as novel therapeutic targets, and several recent reviews highlight the potential of these novel checkpoint targets8,9. The ligands to these receptors can be found, similarly to PD-L1, on the surface of tumor cells and/or antigen presenting cells, meaning that inhibition of TIL via these signaling pathways is expected to occur within tumors. Indeed, patent applications have already been filed around the world for targeting these receptors therapeutically9.
One major reason for the continuing development of novel checkpoint blockade immunotherapies is that only a fraction of patients respond to PD-(L)1 or CTLA4 blockade, and resistance to these therapies can develop in patients that do initially present with a response10,11. Some patients, for example, decrease expression of specific checkpoint markers in a biologic escape mechanism to these therapies, but other molecules with similar cellular functions could still be targeted to achieve similar effects12. As a new wave of immunotherapies makes its way through the pipeline, this is an exciting time to see the growing potential of targeting so many components of the immune system to treat cancer.