References
1. Hagn M, Jahrsdorfer B. 2012. Why do human B cells secrete granzyme B? Insights into a novel B-cell differentiation pathway. 1(8):1368-1375.
2. Tschopp CM, SPiegl N, Didichenko S, Lutmann W, Julius P, Virchow JC, Hack CE, Dahinden CA. 2006. Granzyme B, a novel mediator of allergic inflammation: its induction and release in blood basophils and human asthma. 108(7):2290-2299.
3. Strik MC, de Koning PJ, Kleijmeer MJ, Bladergroen BA, Wolbink AM, Griffith JM, Wouters D, Fukuoka Y, Schwartz LB, Hack CE, van Ham SM, Kummer JA. 2007. Human mast cells produce and release the cytotoxic lymphocyte associated protease granzyme B upon activation. Mol Immunol. 44(14):3462-3472.
4. Wagner C, Iking-Konert C, Denefleh B, Stegmaier S, Hug F, Hansch GM. 2004. Granzyme B and perforin: constitutive expression in human polymorphonuclear neutrophils. 103(3):1099-1104.
5. Hehmann-Titt G, Schiffer S, Berges N, Melmer G, Barth S. 2013. Improving the Therapeutic Potential of Human Granzyme B for Targeted Cancer Therapy. 2:19-49.
6. Hoves S, Trapani JA, Voskoboinik I. 2010. The battlefield of perforin/granzme cell death pathways. J Leukoc Biol. 87(2):237-43.
7. Ewen CL, Kane KP, Bleackley RC. 2012. A quarter century of granzymes. Cell Death Differ. 19(1):28-35.
8. Grossman WJ, Revell PA, Lu ZH, Johnson H, Bredemeyer AJ, Ley TJ. 2003. The orphan granzymes of humans and mice. Opin. Immunol. 15:544-552.
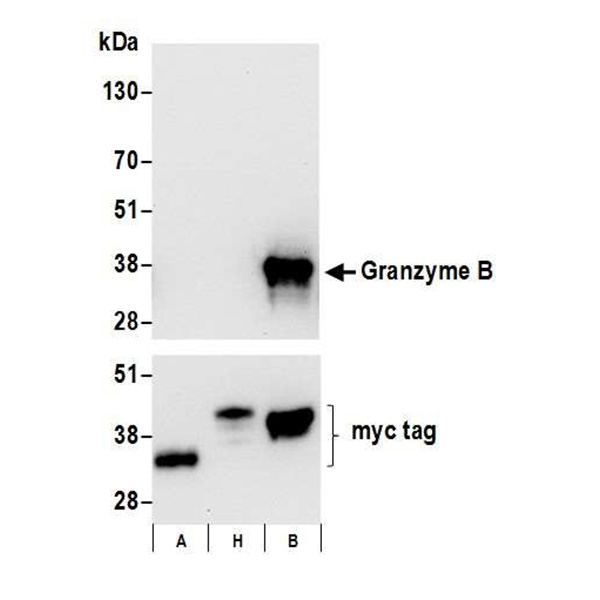
9. Smyth MJ, McGuire MH, Thia KY. 1995. Expression of recombinant human granzyme B. A processing and activation role for dipeptidyl peptidase I. J Immunol. 154(12):6299-6305.
10. Afonina IS, Cullen SP, Martin SJ. 2010. Cytotoxic and non-cytotoxic roles of the CTL/NK protease granzyme B. Immunol Rev. 235(1):105-116.
11. Waterhouse NJ, Sedelies KA, Trapani JA. 2006. Role of Bid-induced mitochondrial outer membrane permeabilization in granzyme B-induced apoptosis. Immunol Cell Biol. 84(1):72-78.
12. Hagn M, Schwesigner E, Ebel V, Sontheimer K, Maier J, Beyer T, Syroets T, Laumonnier Y, Fabricius D, Simmet T, Jahrsdorfer B. 2009. Human B cells secrete granzyme B when recognizing viral antigens in the context of the acute phase cytokine IL-21. J Immunol. 183(3):1838-1845.
13. Holm C, Nyvold CG, Paludan SR, Thomsen AR, Hokland M. 2006. Interleukin-21 mRNA expression during virus infections. 33(1):41-45.
14. Kurschus FC and Jenne DE. 2010. Delivery and therapeutic potential of human granzyme B. Immunol Rev. 235(1):159-171.
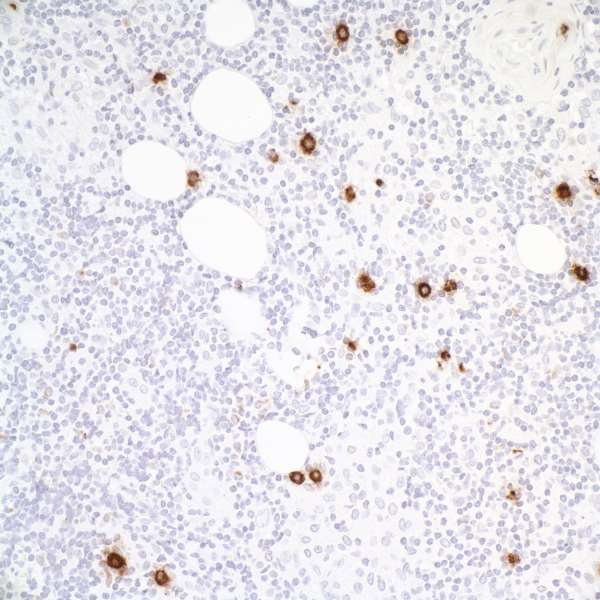
15. Tiacci E, Doring C, Brune V, van Noesel CJ, Klapper W, Mechtersheimer G, Falini B, Kuppers R, Hansmann ML. 2012. Analyzing primary Hodgkin and Reed-Sternberg cells to capture the molecular and cellular pathogenesis of classical Hodgkin lymphoma. 120:4609-4620.